

Which Of Following Is True About The Cytoplasm Of An Animal Oocyte?
In one respect at least, eggs are the most remarkable of animate being cells: once activated, they can requite rise to a complete new individual within a thing of days or weeks. No other cell in a higher animal has this chapters. Activation is usually the consequence of fertilization—fusion of a sperm with the egg. In some organisms, yet, the sperm itself is not strictly required, and an egg can be activated artificially past a variety of nonspecific chemical or physical treatments. Indeed, some organisms, including a few vertebrates such as some lizards, unremarkably reproduce from eggs that get activated in the absence of sperm—that is, parthenogenetically.
Although an egg can give rise to every jail cell blazon in the developed organism, it is itself a highly specialized cell, uniquely equipped for the unmarried office of generating a new individual. The cytoplasm of an egg can even reprogram a somatic cell nucleus and so that the nucleus tin can direct the development of a new individual. That is how the famous sheep Dolly was produced. The nucleus of an unfertilized sheep egg was destroyed and replaced with the nucleus of an adult somatic prison cell. An electric daze was used to actuate the egg, and the resulting embryo was implanted in the uterus of a surrogate mother. The resulting normal adult sheep had the genome of the donor somatic cell and was therefore a clone of the donor sheep.
In this section, we briefly consider some of the specialized features of an egg before discussing how it develops to the bespeak of being prepare for fertilization.
An Egg Is Highly Specialized for Contained Development, with Big Nutrient Reserves and an Elaborate Glaze
The eggs of most animals are giant single cells, containing stockpiles of all the materials needed for initial development of the embryo through to the stage at which the new individual can begin feeding. Before the feeding stage, the giant cell cleaves into many smaller cells, but no net growth occurs. The mammalian embryo is an exception. It can start to grow early on by taking upwards nutrients from the mother via the placenta. Thus, a mammalian egg, although notwithstanding a large cell, does not have to be every bit large as a frog or bird egg, for instance. In general, eggs are typically spherical or ovoid, with a diameter of about 0.1 mm in humans and sea urchins (whose feeding larvae are tiny), 1 mm to ii mm in frogs and fishes, and many centimeters in birds and reptiles (Effigy 20-19). A typical somatic cell, by contrast, has a diameter of only virtually 10 or 20 μm (Effigy twenty-20).

Figure xx-xix
The bodily sizes of iii eggs. The human being egg is 0.1 mm in bore.

Figure 20-20
The relative sizes of various eggs. Sizes are compared with that of a typical somatic cell.
The egg cytoplasm contains nutritional reserves in the form of yolk, which is rich in lipids, proteins, and polysaccharides and is usually independent inside detached structures called yolk granules. In some species, each yolk granule is membrane-enclosed, whereas in others it is not. In eggs that develop into large animals outside the mother'southward body, yolk can account for more than 95% of the volume of the cell. In mammals, whose embryos are largely nourished past their mothers, there is trivial, if whatsoever, yolk.
The egg coat is another peculiarity of eggs. It is a specialized class of extracellular matrix consisting largely of glycoprotein molecules, some secreted by the egg and others deposited on it by surrounding cells. In many species, the major coat is a layer immediately surrounding the egg plasma membrane; in nonmammalian eggs, such equally those of sea urchins or chickens, it is called the vitelline layer, whereas in mammalian eggs it is called the zona pellucida (Figure xx-21). This layer protects the egg from mechanical damage, and in many eggs it also acts every bit a species-specific bulwark to sperm, admitting only those of the aforementioned or closely related species.

Figure 20-21
The zona pellucida. (A) Scanning electron micrograph of a hamster egg, showing the zona pellucida. (B) A scanning electron micrograph of a similar egg in which the zona (to which many sperm are attached) has been peeled dorsum to reveal the underlying plasma (more...)
Many eggs (including those of mammals) contain specialized secretory vesicles just under the plasma membrane in the outer region, or cortex, of the egg cytoplasm. When the egg is activated by a sperm, these cortical granules release their contents by exocytosis; the contents of the granules act to alter the egg coat so as to prevent more than than one sperm from fusing with the egg (discussed below).
Cortical granules are usually distributed evenly throughout the egg cortex, just in some organisms other cytoplasmic components have a strikingly asymmetrical distribution. Some of these localized components later on serve to assist establish the polarity of the embryo, as discussed in Affiliate 21.
Eggs Develop in Stages
A developing egg is called an oocyte. Its differentiation into a mature egg (or ovum) involves a series of changes whose timing is geared to the steps of meiosis in which the germ cells go through their two final, highly specialized divisions. Oocytes have evolved special mechanisms for arresting progress through meiosis: they remain suspended in prophase I for a prolonged period while the oocyte grows in size, and in many cases they subsequently arrest in metaphase Ii while awaiting fertilization (although they can arrest at various other points, depending on the species).
While the details of oocyte evolution (oogenesis) vary from species to species, the general stages are like, as outlined in Figure 20-22. Primordial germ cells migrate to the forming gonad to become oogonia, which proliferate by mitosis for a catamenia before differentiating into primary oocytes. At this stage (usually before nativity in mammals), the first meiotic division begins: the Deoxyribonucleic acid replicates then that each chromosome consists of 2 sister chromatids, the duplicated homologous chromosomes pair along their long axes, and crossing-over occurs betwixt nonsister chromatids of these paired chromosomes. Afterwards these events, the cell remains arrested in prophase of division I of meiosis (in a state equivalent, equally nosotros previously pointed out, to a G2 phase of a mitotic sectionalisation cycle) for a menses lasting from a few days to many years, depending on the species. During this long period (or, in some cases, at the onset of sexual maturity), the primary oocytes synthesize a glaze and cortical granules. In the example of large nonmammalian oocytes, they likewise accumulate ribosomes, yolk, glycogen, lipid, and the mRNA that will later direct the synthesis of proteins required for early embryonic growth and the unfolding of the developmental program. In many oocytes, the intensive biosynthetic activities are reflected in the structure of the chromosomes, which decondense and form lateral loops, taking on a characteristic "lampbrush" appearance, signifying that they are very busily engaged in RNA synthesis (encounter Figures 4-36 and 4-37).

Figure twenty-22
The stages of oogenesis. Oogonia develop from primordial germ cells that migrate into the developing gonad early in embryogenesis. Later a number of mitotic divisions, oogonia brainstorm meiotic partition I, after which they are called primary oocytes. In mammals, (more than...)
The next phase of oocyte development is called oocyte maturation. It ordinarily does not occur until sexual maturity, when the oocyte is stimulated by hormones. Nether these hormonal influences, the jail cell resumes its progress through division I of meiosis. The chromosomes recondense, the nuclear envelope breaks down (this is generally taken to mark the start of maturation), and the replicated homologous chromosomes segregate at anaphase I into 2 daughter nuclei, each containing half the original number of chromosomes. To terminate partitioning I, the cytoplasm divides asymmetrically to produce two cells that differ greatly in size: one is a modest polar body, and the other is a large secondary oocyte, the precursor of the egg. At this stage, each of the chromosomes is still equanimous of ii sister chromatids. These chromatids do non separate until partition II of meiosis, when they are partitioned into separate cells, as previously described. Later on this final chromosome separation at anaphase Two, the cytoplasm of the large secondary oocyte over again divides asymmetrically to produce the mature egg (or ovum) and a 2d small polar body, each with a haploid set of single chromosomes (encounter Figure 20-22). Because of these two asymmetrical divisions of their cytoplasm, oocytes maintain their large size despite undergoing the ii meiotic divisions. Both of the polar bodies are small, and they eventually degenerate.
In well-nigh vertebrates, oocyte maturation proceeds to metaphase of meiosis Ii and then arrests until fertilization. At ovulation, the arrested secondary oocyte is released from the ovary and undergoes a rapid maturation pace that transforms it into an egg that is prepared for fertilization. If fertilization occurs, the egg is stimulated to complete meiosis.
Oocytes Use Special Mechanisms to Grow to Their Big Size
A somatic jail cell with a diameter of x–20 μm typically takes about 24 hours to double its mass in preparation for cell division. At this charge per unit of biosynthesis, such a cell would take a very long time to achieve the thousand-fold greater mass of a mammalian egg with a diameter of 100 μm. Information technology would take even longer to reach the million-fold greater mass of an insect egg with a bore of thousand μm. However some insects alive only a few days and manage to produce eggs with diameters even greater than 1000 μm. It is clear that eggs must take special mechanisms for achieving their big size.
One uncomplicated strategy for rapid growth is to accept extra cistron copies in the cell. Thus, the oocyte delays completion of the first meiotic partitioning so equally to grow while it contains the diploid chromosome gear up in indistinguishable. In this mode, it has twice as much DNA available for RNA synthesis as does an boilerplate somatic cell in the G1 stage of the cell wheel. The oocytes of some species become to even greater lengths to accrue actress Dna: they produce many extra copies of certain genes. We discuss in Chapter half-dozen how the somatic cells of most organisms require 100 to 500 copies of the ribosomal RNA genes in order to produce enough ribosomes for protein synthesis. Eggs crave even greater numbers of ribosomes to back up protein synthesis during early on embryogenesis, and in the oocytes of many animals the ribosomal RNA genes are specifically amplified; some amphibian eggs, for example, incorporate 1 or ii million copies of these genes.
Oocytes may also depend partly on the synthetic activities of other cells for their growth. Yolk, for example, is unremarkably synthesized outside the ovary and imported into the oocyte. In birds, amphibians, and insects, yolk proteins are made past liver cells (or their equivalents), which secrete these proteins into the blood. Inside the ovaries, oocytes take up the yolk proteins from the extracellular fluid by receptor-mediated endocytosis (see Figure 13-41). Nutritive help can too come from neighboring accessory cells in the ovary. These can be of two types. In some invertebrates, some of the progeny of the oogonia go nurse cells instead of becoming oocytes. These cells commonly are connected to the oocyte by cytoplasmic bridges through which macromolecules tin can laissez passer directly into the oocyte cytoplasm (Figure 20-23). For the insect oocyte, the nurse cells industry many of the products—ribosomes, mRNA, protein, then on—that vertebrate oocytes take to manufacture for themselves.
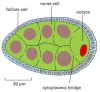
Figure 20-23
Nurse cells and follicle cells associated with a Drosophila oocyte. The nurse cells and the oocyte arise from a mutual oogonium, which gives ascension to ane oocyte and fifteen nurse cells (merely 7 of which are seen in this plane of department). These cells remain (more...)
The other accessory cells in the ovary that help to nourish developing oocytes are ordinary somatic cells called follicle cells, which are institute in both invertebrates and vertebrates. They are arranged equally an epithelial layer effectually the oocyte (Effigy twenty-24, and see Effigy 20-23), to which they are continued only by gap junctions, which allow the commutation of small molecules but not macromolecules. While these cells are unable to provide the oocyte with preformed macromolecules through these communicating junctions, they may help to supply the smaller forerunner molecules from which macromolecules are fabricated. In addition, follicle cells often secrete macromolecules that contribute to the egg glaze, or are taken up by receptor-mediated endocytosis into the growing oocyte, or act on egg cell-surface receptors to control the spatial patterning and centric asymmetries of the egg (discussed in Chapter 21).
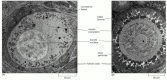
Figure 20-24
Electron micrographs of developing primary oocytes in the rabbit ovary. (A) An early stage of primary oocyte development. Neither a zona pellucida nor cortical granules have developed, and the oocyte is surrounded by a unmarried layer of flattened follicle (more...)
Summary
Eggs develop in stages from primordial germ cells that migrate into the developing gonad early in development to go oogonia. After mitotic proliferation, oogonia become principal oocytes, which brainstorm meiotic division I and then arrest at prophase I for days to years, depending on the species. During this prophase-I arrest period, primary oocytes grow, synthesize a coat, and accumulate ribosomes, mRNAs, and proteins, often enlisting the assistance of other cells, including surrounding accompaniment cells. In the process of maturation, primary oocytes complete meiotic division I to form a small polar body and a big secondary oocyte, which proceeds into metaphase of meiotic division Two. There, in many species, the oocyte is arrested until stimulated by fertilization to consummate meiosis and brainstorm embryonic evolution.
Source: https://www.ncbi.nlm.nih.gov/books/NBK26842/
Posted by: salinasgise1989.blogspot.com
0 Response to "Which Of Following Is True About The Cytoplasm Of An Animal Oocyte?"
Post a Comment